2013年度版 前年も更新しないままになってしまいました。今回も2年ぶりのグループ紹介になってしまっています。 さて、やっと今年度の話に入ります。名黒グループも発足以来9年目に入りました。メンバーはD2渡邊(謙吾)君、D1神山さん、M2椎崎君、丹羽君、M1河原崎君、芝本さん、4年花房君、町田君という8人構成です。昨年度博士を修了した服部君が、石川君と古谷君と一緒に新たなグループとして独立したのと、関根グループからいまだ大型新人の河原崎君、藤澤グループから6年制の芝本さんが入り、野球の得意な(多分実験も)新4年生2人が入ったという動きになりました。
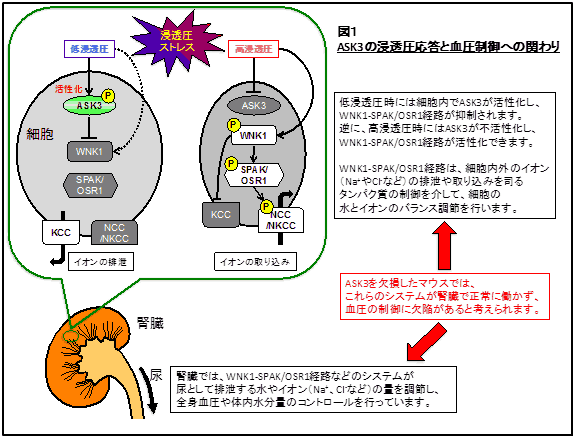
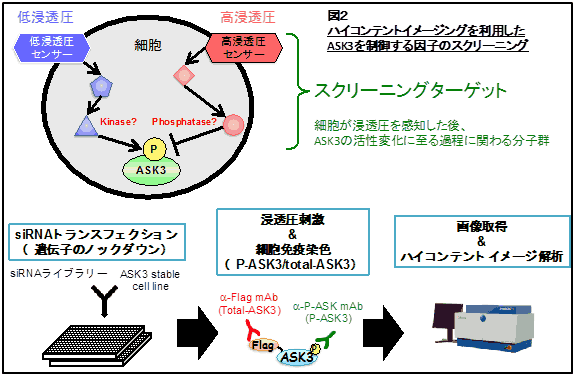
“浸透圧に応じて”と書きましたが、ASK3の浸透圧ストレスに対する応答性は他の分子にはあまり見られない独自のものです。以前にも書いたように、ASK3は細胞の周囲が低浸透圧になればなるほど活性化し、高浸透圧になればなるほど不活性化するという連続的で、かつ両方向性の活性変化を示します。応答の早さも1-2分で変化する素早いもので、可逆的であることから、ASK3は細胞が周囲の浸透圧変化を測るための浸透圧メーターのような働きをしていると考えられます。また、ASK3は『浸透圧という物理的な量』を『キナーゼ活性によるリン酸化反応の程度』という細胞内の化学的な反応に変換する変換機として働いていると見ることもできます。生体のストレス応答とは、生存に適した状態から環境がシフトした場合に、それに対応して適応しようとする動きです。その際、『シフトの方向と程度を測り、それに応じたアウトプットを出力する』ことが必須項目であり、まさにASK3は浸透圧ストレス応答において適した性質を持つ分子だと考えられます。 さて、ここで、ASK3の浸透圧に依存する独特の活性変化がどのようにして引き起こされているかという疑問が浮かびます。実は、そもそもヒトの細胞自体が浸透圧をどうやって感知しているかについて未だ不明な点が多く残されています。この問題について、私たちはASK3の活性変化を指標にしてアプローチしようと考えています。ASK3はそれ自身の活性がリン酸化により制御されているため、浸透圧依存的にASK3のリン酸化を制御する分子が存在すると考えられます(図2)。
この他にも、ASK3が関わるシグナル伝達経路の解明のために、分子特異的クロスリンク法という独創的なケミカルバイオロジーを利用したASK3結合分子の探索と解析や、ASK3ノックアウトマウスを用いて、in vivoレベルでの水・イオンバランスの恒常性維持にASK3が関与するかなどに関する解析も行っています。最近、高食塩食が高血圧を引き起こすだけでなく、免疫細胞の活性化を介して自己免疫疾患の程度を悪化させるという論文なども報告されており、浸透圧やイオンバランスの維持が慢性疾患をはじめとする様々な病態の理解に重要であると考えられます。私たちのASK3を一つの手がかりとして生体の浸透圧ストレス応答機構を解明しようとする研究は、このような病態の理解や新たな治療法の確立に貢献する可能性を秘めていると思っています。また、2011年度に書いたように、個人的には浸透圧ストレスだけでなく、温度ストレスやメカノストレスなど両方向性に変化する刺激に対して、生体がどのように応答するかについても興味があり、今後さらに解析を進めていきたいと思っています。 名黒グループには、昨年度、今年度とグループの再編に伴い、他のグループから新たに参加した大学院生もおり、彼らの持つ知識や技術を活かしたテーマも行っています。具体的には、がん転移におけるASKファミリーの役割の解析や、センチュウを利用したシグナル伝達経路の遺伝学的解析です。 名黒グループも9年目になり、私自身もアラフォーに手がかかる歳に達しました。それに見合うだけの働きと、存在感を持たなければと思う今日この頃です。しかし、研究生活のなかで、新しい人、新しい技術、新しい実験データが与えてくれるインパクトを見ていると、変に落ち着いてはダメで、いつも新しいものを求める気持ちを持ち続けようと思います。毎日得られる新しいことを周りの人達、グループ員と共有しながら研究生活を進めていきたいと思っています。 |
|
||||||||
2011年度版 またもや、昨年度版を出さないまま、2011年度を迎えてしまいました。ごめんなさい。 |
近年、メカノストレスは筋肉や骨の発達のみならず、がんの悪性化、ES細胞の分化方向決定など様々な生命現象に深く関わっていることが明らかになりつつあり、盛んに研究されている分野でもあります。これまでにメカノストレスに対する応答では、アクチン、ミオシン、チュブリンなどをはじめとした細胞骨格系タンパクや、細胞の外界との接着点である接着斑(focal adhesion)などに局在するタンパクの関与が数多く報告されています。しかし、これまでの研究でASK3の浸透圧依存的な活性変化はこれらの分子の働きを必要とせず、ある程度独立して起きている現象らしいという結果を得ており、新たなメカノストレス応答のシステムに含まれている可能性があります。今後、細胞膜や膜タンパクとの相互作用などを検討しつつ、ASK3がどのような状況のメカノストレスで働くか研究していきたいと考えています。 ともすれば、研究とはなにやら難しい世界の話と捉えがちですが、浸透圧、温度、物理的な刺激などは自分たちの周りにいくらでもあるありふれた日常の両方向性のストレスであり、誰でも知らず知らずに応答し適応しているものです。毎日自分の手のひらの上で起こっているような、身近で且つ大切な生命現象の源を解き明かしていくこと自体がとても面白いことだと思います。また同時に、そのような身近な生命現象の不調や破綻が、実は様々な病態の原因に関わっているのではないかと考え、研究を行っています。 |
|||||||
|
||||||||
2009年度版 2009年度になり、とうとう名黒グループも5年目になりました。 グループの研究テーマとしてはASK3というリン酸化酵素についての研究を継続しています(詳細は前年度以前を参照)。ASK3は低浸透圧に対して活性化、高浸透圧に対して不活性化するという非常に面白い“両方向性の”キナーゼ活性の変化を示しますが、この独特の浸透圧ストレスに対する活性変化がどのような分子メカニズムにより達成されているかについては全く分かっていません。私たちの研究からASK3に結合するチロシンキナーゼやASK3を脱リン酸化するフォスファターゼが明らかになり、これらによるASK3の活性制御機構を現在解明しようとしているところです。 また、ASK3活性制御メカニズムを担う分子群を網羅的に同定するために今年からひとつ新しい技術を導入しました。High-Content Analysis(HCA)技術と呼ばれる大規模なスクリーニングなどを行うためのスループットに優れたシステムです。今回導入したのは薬学部の共通機器室に設置されたThermofisher社のArrayScan VTIという画像解析機器で、この機械は細胞一つ一つの情報を数千から数万個という単位で短時間に集めることができます。この技術のすごさを説明するために、例えば私たちがASK3の解析で行ってきたリン酸化抗体を利用したキナーゼ活性の測定について、これまでのウェスタンブロットと比較してみます。 |
ただ、これまでは細胞免疫染色画像からこの数十万という数を解析することが現実的に無理でした。蛍光顕微鏡で個々の細胞について人がピントを合わせて測定し、次の細胞にいくなどということをすると時間がいくらあっても足りません。この部分を機械が細胞を見分け、サンプル中にある全ての細胞について即座に解析を行うことができるようにしているのがArrayScanという機器です。これがHCAたる所以です。 先に触れたHCAなどの新しい技術や方法論が開発されるに従って、生物を学問として解明していくやり方もどんどん増えていきます。もちろんこれらの技術を貪欲に取り入れて研究を進めていきたいと思います。ただ、それでもやはり技術革新に頼りきるのではなく、自分たちの頭からひらめく新しいアイディアも武器として、さらに生き物を読み解いていく研究を進めていきたいと考えています。 |
|||||||
2008年度版 2008年度になり、名黒グループも4年目に入りました。昨年度は紅一点だった高木さんが修士過程を修了して卒業して、男だらけになる可能性があったのですが、新4年生加入によりなんとかその危機は回避されました。新4年生は、小出さん、清水君の2人が加わり、昨年下積みをつんだ橋本君、服部君がしっかり指導をしてくれています(稀にたじたじのことも…)。また、その上にはM2になり、(いい意味で?)ますます目つきの鋭くなった丸山君が控えており、全体を見渡す位置にD2の梅田君が鎮座ましましている(さぼっている訳ではありません)というのが名黒グループの現状です。合計で院生・学生は6人になりました。なかなか賑やかです。 新しく入ってきた4年の2人も興味を持って研究を始めているようですし、もちろん、もとからいた人達も新しいことを発見しつつ、妄想(昨年度参照)を膨らませて研究を進めていってくれています。グループヘッド当人はとうとう三十路に入り、若さが武器っ!とはおおっぴらには言えなくなってきた感じですが、でも、まだまだがんばります! さて、グループの研究テーマとしては、以前から継続してASK3という新しく同定したリン酸化酵素の制御と生理的役割の解析を行っています。 生物学において、リン酸化酵素など単独の分子をテーマとするといった場合、生き物を個体→臓器→組織→細胞→分子というような順で解体(微分)していくと理解し易い、という分子生物学的視点が前提となっています。従って、実験で分子について得られた知識は、その逆の変換(積分)により生き物の個体レベルの現象を説明する知識にしていきたいというのが、個人的な持論です。 これを適用すると、うちのグループの大きな目的の一つは“ヒトは何のためにASK3を持っているのか?”ということを分かりやすく解明することです。言い換えれば、これまで体の中で知らず知らずのうちに働いていたASK3という分子の役割を人間に分かる形で明らかにしてやろう!ということです。ASK3は生理的役割についてまだ全く報告のない分子なので、とても面白いテーマだと思っています。 とはいえ、この分子の知識から個体の現象を説明しようとする“積分”はとても難しいもの。まずは分子自体の性質を詳しく知らなければ手も足もでません。そうして分子の解析をしてみると、生命現象の小さな単位としての分子自体も研究対象として複雑でなかなか面白いものだということが分かります。 さらに、ASK3の生理的役割を知るためには当然生体内の他の分子との関わりを考える必要が出てきて、ASK3の研究をしているつもりでも結局は様々な分子群が絡む細胞内の情報伝達経路が研究の対象になってきます。研究をすればするほど、研究対象として扱う世界が広がっていきます。その延長線上に個体におけるASK3の役割や、病態への関与、薬のターゲットとしての可能性などが見えてくると考えています。 抽象的な話はこの辺りにして、具体的にうちのグループで行っている研究内容を以下に記します。 |
リン酸化酵素(分子)としてのASK3の解析 ASK3の担う細胞内シグナル伝達経路の解析 個体におけるASK3の役割の解析 沸き上がる数多くの妄想を検証すべく、グループ全体一丸となって日々研究を行っています。 |
|||||||
|
||||||||
2007年度版 2006年度版はただいま作成中という状態のまま、2007年度になってしまいました。すみません。昨年度も永田くんと丸山君というでかい二人が入ったり、河野君が修士を取得して卒業したりといろいろあったのですが、、、 開き直って2007年度版のグループ紹介をします。 名黒グループも3年目に入り、メンバーも少し増えて6人になりました。初めての博士課程(梅田君 D1)、修士課程(高木さんM2、丸山君 M1)、4年生(橋本君、服部君)とバリエーションに富んだ人員ですが、平均するとうちのグループの中で一番若い。一応グループヘッドも20代最後の年。ということでまだまだ発展途上ですが、勢い(内側で燃やす系です。【注】丸山君は外も)だけは負けないぞ!というグループです。 グループの研究テーマとしては、研究室自体のテーマであるストレス応答に関与する細胞内シグナル伝達の解析のうち、ASK3という新しく同定したリン酸化酵素の制御と生理的役割の解析を行っています。これまでの解析でこの分子は物理化学的ストレスの一つである浸透圧により非常に面白い活性変化を示すことが分かってきています。高浸透圧と低浸透圧で異なる変化を示すのです。 物理化学的ストレスには、紫外線、重力、活性酸素、浸透圧、温度など様々ありますが、これらのうち浸透圧、温度、pHなどは生き物にとって至適条件のある量です。つまり浸透圧なら約300 mOsm/kg H2Oという正常な値があり、それを上回っても下回っても生体にとってストレスになります。これらのストレスに対しては上がれば下げて、下がれば上げてという両方向性の制御があって初めて正常に保てることになります。 名黒グループではASK3というストレス変化の方向性依存的に異なる応答を示す分子を独特な切り口として、浸透圧のような両方向性の制御を必要とする量の細胞レベルでの受容と応答のメカニズムについて解明していきたいと考えています。そして、それは個体レベルでのストレス応答を理解する上で大きな手がかりになると考えています。 例えば、飲水や過剰な食塩摂取により個体レベルで水やイオン、つまりは浸透圧の恒常性が乱れた場合、うまくストレス応答ができないと体には浮腫や血圧の変化が起こります。血圧は上がりすぎても下がりすぎても病気であり、正常範囲におさめるメカニズムが生体には存在します。それが生命の最小単位である細胞に存在するタンパク質によって担われていると考えられるわけですが、浸透圧変化に応答して活性を変えるASK3はこのようなメカニズムを担うタンパク質の一つではないかと考え解析しているところです。このようにストレス応答に関わる分子の役割を解析することで、高血圧など様々な疾患に対する薬剤の新たなターゲット(分子、メカニズム)を提供できるのではないかと考えています。
|
具体的な研究内容
1)浸透圧変化によるASK3の活性変化がどのような分子基盤により達成されているかについて、ASK3自身の分子構造の観点、活性を制御する結合分子のスクリーニング、さらにはショウジョウバエのRNAiスクリーニング系を用いた遺伝学的手法により解析しています。分子構造の解析ではASK3の独特な活性変化に必要な部分をキメラ分子の作成などにより同定することで、活性制御の構造的基盤が明らかになるものと考えています。結合分子による制御についても様々なスクリーニングの結果数多くの結合分子が見つかり、今後の解析が楽しみです。また、遺伝学的解析は結合分子の探索より広く関連する分子を探索できる手法として、系が順調に立ち上がりつつあります。これらの解析によりASK3がどのようにして浸透圧変化を感受し、リン酸化活性に変換しているか明らかにしていきたいと考えています。 2)ASK3が担う細胞内のリン酸化シグナル伝達経路についてもいろいろと分かってきています。結合分子として得られてきた遺伝性高血圧の原因分子がASK3のリン酸化による制御を受けていそうだということが分かり、現在このシグナル伝達経路に注目して解析しています。先に書いたようにASK3が実際に個体レベルの血圧制御に関与している可能性もあるのではないかと考えています。また、多くのストレスで活性化されるMAPKの一つが浸透圧ストレスにおいてASK3の下流にあることも明らかになり、この意義についてもイオンの輸送や遺伝子発現などの観点から解析を行っています。 3)分子や細胞レベルの解析によるASK3の知見を個体レベルにおけるASK3の役割に還元するため、各組織での発現分布の解析、発現臓器に由来するcell lineの利用、ASK3ノックアウトマウスの作成なども行っています。現在、もうすぐノックアウトマウスが誕生するという段階まできており、生まれるのを心待ちにしている状態です。このノックアウトマウスの解析などによりASK3が生体のどのような機能に重要な役割を果たしているか、病態との関連などはあるのかについて明らかにできるものと考えています。 この他にも、タンパク質の解析に欠かせないモノクローナル抗体の作成も行っています。良質な抗体は実験の幅を広げてくれる大切なツールであり、それを自作できることは強力な武器だと考えています。実際に当教室で使用しているASK3の抗体はグループ内で作成したとても良いものです。現在も解析したい分子の抗体やリン酸化特異的抗体の作成などをどんどん進めており、モノクロ抗体作成のノウハウは業者にも劣らないものと自負しています。 名黒グループができてASK3の解析を始めた頃に比べると様々な点が明らかになってきており、この分子の機能や役割について仮説(妄想ともいう)を膨らませる楽しさも倍増してきました。また、一つの疑問に答えが出ると、新たな謎が現れるという繰り返しでもあるので、グループメンバーはみんな忙しく実験しています。思いついた一つ一つの仮説を実験によって肯定、もしくは否定(こちらのほうが難しい)して、妄想を事実に近づけていくために日々研究を行っています。 |
|||||||
| 2005年度版
名黒サブグループ 名黒サブグループは2005年4月に名黒 功(なぐろ いさお)が細胞情報学教室の助手として就任するに当たり発足したグループです。 私、名黒は学生時代、現在の細胞情報学の前身である薬効安全性学教室という薬理学の研究室でカルシウムチャネルの電気生理を中心に学んでいました。博士課程に在籍中、現在の一條教授の細胞情報学教室に教室が変わり、細胞生物学という新たな領域にふれる機会がありました。この生理学、病理学的視点を色濃く持つ薬理学の教室と、分子レベルの生化学を扱うことのできる細胞生物学の教室、両方を経験できたことは自分にとってとても大きなものだったと考えています。薬学の一つの目的である基礎研究の医療への還元は、個体の病気の症状や病態を正しく把握し、それを分子レベルまで分解して理解することで初めて薬という形を取ると考えられます。その課程で大切な病態を把握する生理学的知識と、現象を分子レベルまで分解する生化学的知識の両端に触れられた経験を生かしていきたいと考えています。つまり、うちのグループでは生理学、薬理学的視点を活用しながら分子レベルまでの解析を用いて生体機構、病理機構を解明する研究を行っていきたいと思っています。 現在、当グループの研究テーマは、新たに発見したあるリン酸化酵素の解析を中心に据えて行っています。この分子に関してはまだまだ未知の部分が多く、やりたいことがいっぱいあります。 |
その最たるもので現在取り組んでいるのがこの分子のノックアウトマウス作成です。これまでに、このリン酸化酵素が腎臓に多く発現し、ストレスの一つである浸透圧の変化によって活性を変化させることが分かりました。親グループである松沢グループにも書いてありますが、細胞内の分子の現象を個体レベルの表現系と関連づけて考えるために大きな助けとなるのが、ノックアウトマウスの解析です。ノックアウトマウスができればこのリン酸化酵素の生理的役割や病態への関与について明らかになっていくはずです。
リン酸化酵素の役割の理解には、活性化状態と不活性化状態を単に行き来する分子という認識だけでなく、その分子がいつ(活性化刺激、発達段階)、どこで(組織、細胞内局在)、どれくらい(活性化程度と持続時間)活性化するかという知見が大切です。これらを把握することは生命現象を一度分解して得た一つの分子の働きを個体の生理反応に正しく再構成するうえで必須です。そのため、従来当研究室で洗練、蓄積されてきたタンパクを扱う手法に加えて、細かい時間、空間分解能を持つ共焦点レーザー顕微鏡などの手法も用いてこの分子の性質の解析を行っています。 さらに、この分子の基質および活性を制御する分子の同定を目的に、浸透圧依存的(リン酸化活性依存的)に結合する分子の探索も行っています。このような分子の同定から、このリン酸化酵素が担うシグナル伝達経路がどのような生理応答に関与するか、その位置づけが明らかになると期待されます。 名黒グループの研究はまだ始まったばかりでどのように発展していくか未知な部分も多いですが、だからこそいろいろな驚きと感動を経験できると考えて日々の研究を行っています。 名黒グループの研究内容について興味のある方、またはこのページに対してのご意見やご感想は以下のメールアドレスにお気軽にメールしてください。 nagurois@mol.f.u-tokyo.ac.jp |
|||||||